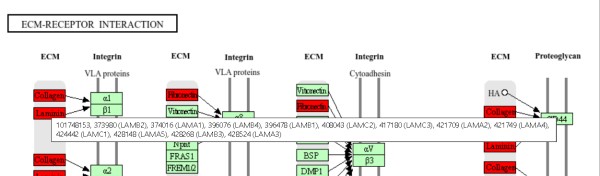
细胞外基质(ECM)与细胞之间的相互作用在调节细胞表型和行为方面发挥着至关重要的作用。这些相互作用主要通过细胞表面受体(就是后文的整联蛋白integrins)进行,包括直接或间接与协作分子介导的相互作用。ECM不断地重塑,影响细胞信号通路。ECM大分子的片段化进一步增加了细胞环境的复杂性。当这些相互作用保持平衡时,身体保持健康;相反,这些相互作用的失调可能导致病理过程,最终导致各种疾病。因此,基于保持正常的ECM-细胞相互作用的疗法应用是高度合理的。此外,根据当前的知识,未来开发有效的药物疗法也需要同时针对复杂的ECM-细胞相互作用。
细胞外基质(ECM)是细胞外环境中的一种复杂网络,主要由蛋白质和多糖组成。它存在于所有动物组织中,为细胞提供结构支持和物理锚定,同时也参与调节细胞的生物学行为,如生长、分化和迁移等。ECM的主要成分包括胶原蛋白、弹性蛋白、纤维蛋白、蛋白聚糖和粘连糖蛋白等。这些分子通过相互作用形成三维网络,为细胞提供适宜的微环境,并参与细胞间的通讯和信号传递。动物细胞感知、粘附和重塑局部ECM的能力对于控制细胞形状、机械响应、运动和信号传导至关重要,因此对于发育、组织形成、伤口愈合和免疫反应都是核心。细胞与ECM之间的相互作用发生在各种专门的多蛋白粘附复合物中,这些复合物将ECM物理连接到细胞骨架和细胞内信号装置。这主要发生在整联蛋白家族的聚集跨膜受体上。
在人类中,特定的细胞与ECM之间的相互作用主要由跨膜分子介导,主要是整联蛋白(integrins)以及可能的蛋白聚糖(proteoglycans)、CD36或其他细胞表面相关组分。这些相互作用直接或间接地控制着诸如粘附、迁移、分化、增殖和凋亡等细胞活动。此外,整联蛋白作为机械受体,提供了ECM与细胞骨架之间的力传递物理连接。整联蛋白是一类糖基化的、异二聚体跨膜粘附受体,由非共价结合的α和β亚单位组成(也就是图中的α、β受体,二者是一个整体,构成一个蛋白,这也说明在这个图中,如果两个条框合并在一起表明两个都是一个完整蛋白的亚基,只有合并在一起才能成为一个整体发挥功能,换句话说,这里α、β受体中的一个基因的转录量增加是毫无意义的,因为α和β亚单位是由不同的基因分别表达后组装而成的,某一个亚基的转录量或者表达量增加不会改变最后integrins的表达量,因此除非两个亚基表达量都增加,否则这里可以直接忽略掉integrins的表达情况,从本图来看是可以忽略掉的)。
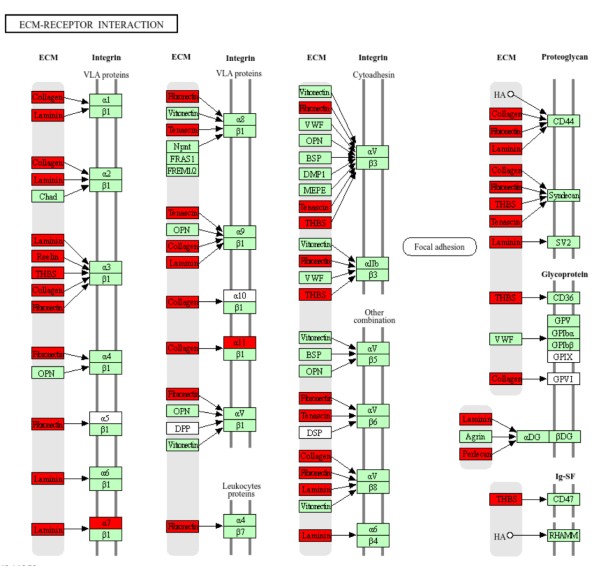
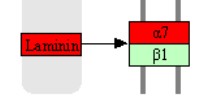
但这并不是说整联蛋白作为受体蛋白不重要,而是说在考虑转录量增加的时候可以不考虑这个问题,但如果涉及表达量下降的话,它就是非常值得关注的信息了,因为任何一个亚基的表达量下降都会带来终点蛋白受体的减少,而受体蛋白减少可能意味着该功能的抑制,比如图中的Integrin α7β1表达量如果下降,可以显著增加肠道细胞的分化和极化标记物的水平,以及形态特征,简而言之就是促进隐窝中的干细胞分化为肠上皮细胞,这样的话就值得关注了。当然,你这个是表达量增加,作用就可能是相反的。
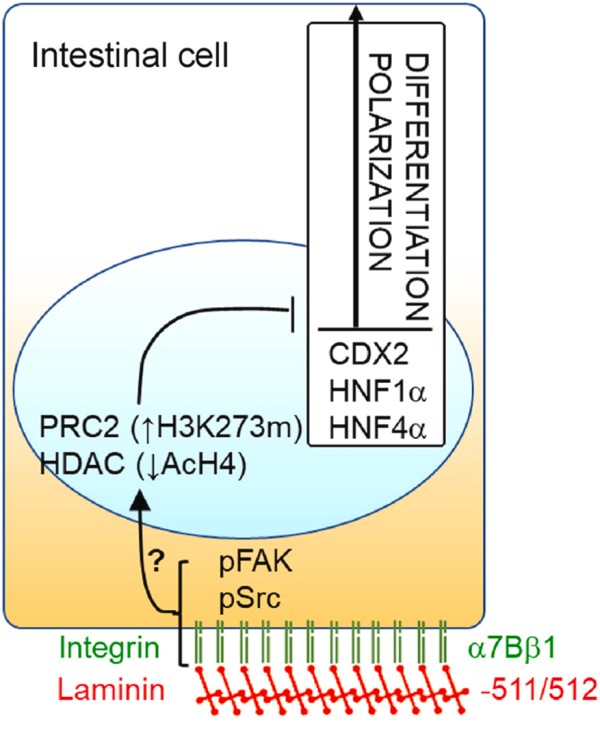
二、通路中表达量增加的ECM
要研究信号通路,一定要搞清楚信号因子到底代表什么?一个基因之所以能够行使多种功能,在于基因转录过程中的关键步骤,即“剪切”,在基因表达过程中,将前体mRNA(pre-mRNA)中的内含子去除,并将外显子序列连接起来,形成成熟的mRNA,进而指导蛋白质的合成。在这个过程中不同的“剪切体”最后形成不同蛋白质,行使不同的功能,这是我们理解信号通路图的基础。
为什么一样是VLA protein却有这么多个不同,正是因为他们对应了α、β亚基不同的剪切体翻译出的蛋白的组合,从而行使不同功能。而KEGG的分析似乎是计算亚基片段的组合可能性来生成信号通路图,